


|
Synonym(s)
Basionym
The original name for an organism. In botany, the original published nomenclature from which a new binomial nomenclature is derived for a particular group of organisms (Tindall 1999).
(basionym) (Guiry 2011)Classification
(Guiry and Guiry 2011)
Lifestyle
Auxospore
In diatoms, the special cells that restore normal size following cell division. Auxospores are associated with sexual reproduction (Horner 2002).
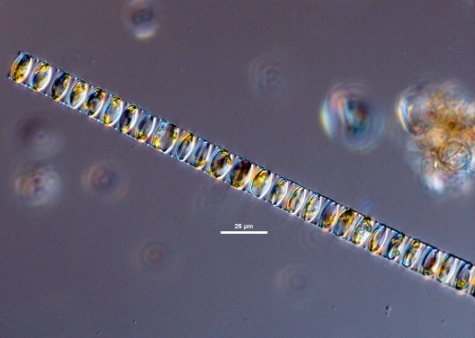
Auxospores are sometimes present (Cupp 1943). Resting stage can survive up to 6 years (Lewis 1999).Description
Margin
The outline or border that defines the shape of an organism or cell.
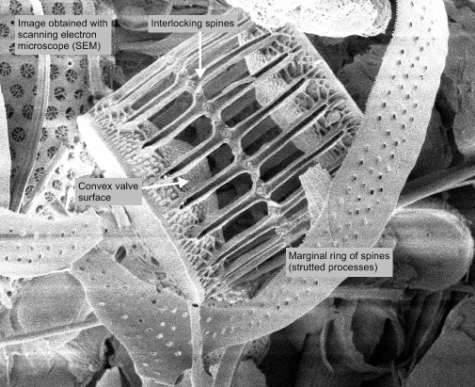
marginal ring of spines CloseStrutted process
(how is this different than marginal process?) In some diatoms, a narrow tube through the frustule that is usually associated with the secretion of chitin. It may appear as a marginal process or as a simple pore in the valve wall (Spaulding et al. 2010).
(strutted processes). The CloseValve
In diatoms, the structurally distinct halves of the cell wall (Becker 1996).
valve face is convex to flat. Spines interlock midway between adjacent cells, visible as a dotted ring; spine lengths are variable (Horner 2002). There are two chloroplasts per cell and the CloseNucleus
(plural: nuclei) In eukaryotic cells, a membrane-bound organelle that contains the cell's genetic information; the nucleus controls the activities of the cell by controlling gene expression.
nucleus is located centrally (Cupp 1943).Labiate process
In diatoms, a simple slit in the valve wall with two internal lips, one on each side of the slit. They can be useful in identification because they are positioned differently in different species (Horner 2002).
labiate process is present near the center of the valve inside the ring of CloseProcess
A natural projection or appendage on an organism.
processes (Horner 2002).Measurements
Pervalvar axis
The axis through the centre point of the two valves of a frustule. This axis is perpendicular to the valve face.
(pervalvar axis): 2 - 61 μmDiameter: 2 - 21 μm
Close
Marginal process
In some diatoms, a long, coarse external tube through the frustule (Tomas 1997).
Marginal processes: 6 - 30 (total)(Hasle and Syvertsen 1997)
Similar species
Harmful effects
Habitat
Distribution
Cosmopolitan in coastal except polar seas (Horner 2002).
Often forms extensive spring blooms (Horner 2002).
One of the most abundant species, especially from February to April (Cupp 1943). Also forms blooms in autumn.
Growth conditions
Irradiance
Amount of solar energy per unit area on a surface (units: μE m-2 sec-1, where E is an Einstein, a mole of photons).
irradiance of 1.6×1016 quanta/s⋅cm2 (Yan et al. 2002). It has an optimal salinity range of 18 - 25 (Yan et al. 2002), although it is capable of significant growth in salinities 5 - 40 (Brand 1984). It is often dominant in waters characterized by salinity fluctuations (Rijstenbil 1987, cited in Rijstenbil 1988).Nutrients
Various chemical substances that an organism needs for metabolism (i.e., to live and grow). These are usually taken up from the environment. Some examples include nitrate, phosphate, silica (for diatoms), iron, copper, etc. Some nutrients, like copper, are required for growth, but can also be toxic at high levels.
nutrients (NO3-, PO43- and CloseSilicic acid
A general term to describe chemical compounds containing silicon, oxygen and hydrogen with a general formula of [SiOx(OH)4-2x]n. Diatoms polymerize silicic acid into biogenic silica to form their frustules (Azam and Chisholm 1976).
silicate) conditions and often forms dense blooms, especially when silicate concentrations are higher than 2 μmol L-1 (Hu et al. 2011). It is also tolerant of CloseEutrophic/eutrophication
Water that is enriched in natural or artificial mineral and organic matter, which promotes an abundance of plant life (i.e., algae), and can result in reduced oxygen conditions.
eutrophication conditions (Abdalla et al. 1995).Environmental Ranges
Temperature range (°C): -1.541 - 29.468
Nitrate (μmol L-1): 0.053 - 28.280
Salinity: 19.590 - 37.775
Oxygen (mL L-1): 4.444 - 9.002
Phosphate (μmol L-1): 0.046 - 2.337
Silicate (μmol L-1): 0.648 - 59.039
(OBIS 2011, cited in EOL 2011)
Bloom characteristics
References
Abdalla, R. R., Zaghloul, F. A. and Hussein, N. R. 1995. A statistical modelling of phytoplankton eutrophication in the Eastern Harbour, Alexandria, Egypt. Bulletin of the National Institute of Oceanography and Fisheries (Egypt). 21(1): 125-146.
Andreae, M. O. and Klumpp, D. 1979. Biosynthesis and release of organoarsenic compounds by marine algae. Environmental Science and Technology, 13, 738�741.
Brand, L. E. 1984. The salinity tolerance of forty-six marine phytoplankton isolates. Estuarine, Coastal and Shelf Science, 18(5): 543-556.
Borkman, D. G. and Smayda, T. 2009. Multidecadal (1959 - 1997) changes in Skeletonema abundance and seasonal bloom patterns in Narragansett Bay, Rhode Island, USA. Journal of Sea Research, 61: 84-94.
Cleve, P. T. 1873. Examination of diatoms found on the surface of the Sea of Java. Bihang till Kongliga Svenska Vetenskaps-Akademiens Handlingar,�1(11): 1-13.
Cupp, E. E. 1943. Marine Plankton Diatoms of the West Coast of North America. University of California Press. Berkeley, California. 238.
Encyclopedia of Life (EOL). Skeletonema costatum�(Greville) Cleve. http://www.eol.org/pages/910783. Accessed 10 Jun 2011.
Guiry, M. D. 2011. Skeletonema costatum�(Greville) Cleve, 1873. http://www.marinespecies.org/aphia.php?p=taxdetails&id=149074. Accessed 05 May 2011.
Guiry, M. D. and Guiry, G. M. 2011. Skeletonema costatum (Greville) Cleve. http://www.algaebase.org/search/species/detail/?species_id=39687. Accessed 05 May 2011.
Hasle, G. R. and Syvertsen, E. E. 1997. Marine diatoms. In: Tomas, C. R. (ed.) Identifying marine Phytoplankon. Academic Press, Inc., San Diego. 5-385.
Hori, Y., Miyahara, K., Nagai, S., Tsujino, K., Nakajima, M., Yamamoto, K., Yoshida, Y., Araki, N. and Sakai, Y. 1998. Relationships between the dominant phytoplankton and DIN:DIP ratios in Osaka Bay and Harima-Nada. Nippon Suisan Gakkaishi. 64(2): 243-248.
Horner, R. A. 2002. A Taxonomic Guide To Some Common Phytoplankton. Biopress Limited, Dorset Press, Dorchester, UK. 200.
Howard, A. G., Comber, S. D. W., Kifle, D., Antai, E. E. and Purdie, D. A. 1995. Arsenic speciation and seasonal changes in nutrient availability and micro-plankton abundance in Southampton Water, U. K. Estuarine, Coastal and Shelf Science, 40(4): 435-450.
Hu, H., Zhang, J. and Chen, W. 2011. Competition of bloom-forming marine phytoplankton at low nutrient concentrations. Journal of Environmental Science, 23(40): 656-663.
Kraberg, A., Baumann, M. and Durselen, C. D. 2010. Coastal Phytoplankton: Photo Guide for Northern European Seas. Verlag Dr. Friedrich Pfeil, Munchen, Germany. 204.
Kuylenstierna, M. and Karlson, B. 2006. Swedish Meteorological and Hydrological Institute: Skeletonema costatum (Greville) Cleve 1873. http://www.smhi.se/oceanografi/oce_info_data/plankton_checklist/diatoms/skeletonema_costatum.htm. Accessed 05 May 2011.
Lewis, J., Harris, A. S. D., Jones, K. J. and Edmonds, R. L. 1999. Long-term survival of marine planktonic diatoms and dinoflagellates in stored sediment samples. Journal of Plankton Research, 20(2): 343-354.
Miralto, A., Barone, G., Romano, G., Poulet, S. A., Ianora, A., Russo, G. L., Buttino, I., Mazzarella, G., Laabir, M., Cabrini, M. and Giacobbe, M. G. 1999. The insidious effect of diatoms on copepod reproduction. Nature. 402: 173-176.
Ocean Biogeographic Information System (OBIS). Skeletonema costatum. http://www.iobis.org/mapper/?taxon_id=510866. Accessed 10 Jun 2011.
Red-Tide. Skeletonema costatum. http://www.red-tide.org/new_site/sc.htm. Accessed 05 May 2011.
Rijstenbil, J. W. 1987. Phytoplankton composition of stagnant and tidal ecosystems in relation to salinity, nutrients, light and turbulence. Netherlands Journal of Sea Research, 21: 113-123.
Rijstenbil, J. W. 1988. Selection of phytoplankton species in culture by gradual salinity changes. Netherlands Journal of Sea Research, 22:�291-300.
Sanders, J. G. and Windom, H. L. 1980. The uptake and reduction of arsenic species by marine algae. Estuarine, Coastal and Marine Science, 10: 555-567.
Sarno, D., Kooistra, W. H. C. F., Medlin, L. K., Percopo, I. and Zingone, A. 2005. Diversity in the genus Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of S. costatum-like species with the description of four new species. Journal of Phycology, 41(1): 151-176.
Yan, T., Zhou, M. and Qian, P. 2002. Combined effects of temperature, irradiance and salinity on growth of diatom Skeletonema costatum. Chinese Journal of Oceanology and Limnology, 20(3): 237-243.
Zingone, A., Percopo, I., Sims, P. A. and Sarno, D. 2005. Diversity in the genus Skeletonema (Bacillariophyceae). I. A re-examination of the type material of S. costatum with the description of S. grevillei sp. nov. Journal of Phycology, 41(1): 140-150.